Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
5.1.9.I Replication Process
I1. Replicative Influence. Can the replicator influence the probability that its own successful replication will occur? (Section 5.1.1)
I2. Replicator Activity/Passivity.
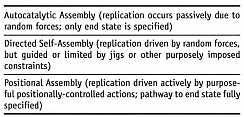
Does the replication process occur probabilistically (stochastically) or deterministically?
For example, enzymes depend on random diffusion events, whereas molecular mills
and nanomanipulator arms operating in a vacuum environment demand positionally-precise
actively-controlled tool tips.
I3. Replicator Parasiticity.
Viral or parasitic character of the device, defined along a continuum; alternatively,
the number of “vitamin processes” (processes not accessible to the
replicator internally), ranging from no vitamin processes to all vitamin processes.
Extreme specialization for replication in a single particular external environment
allows a replicator to eliminate many otherwise essential subsystems, thus making
that replicator highly dependent on the chosen substrate and vulnerable to failure
if the substrate is later altered in any way. Taking natural human endoparasites
as an interesting biological analogy, Knutson [2542]
notes that “when a parasitic creature takes up a serious parasitic lifestyle,
it is likely to lose some important things. It loses its external skin (epidermis)
because it has to be able to absorb food through its body surface. It loses
most of its nervous system, just as creatures that live in the darkness of caves
for thousands of generations are likely to lose their eyes. It loses its digestive
system because all food is conveniently predigested by the host. And it may
lose its capacity to move in favor of a capacity to just hang on....Adult tapeworms
exemplify the extreme losses suffered by parasites. Their bodies are almost
totally dedicated to the development and use of their reproductive apparatus.
They lack digestive systems, since they mostly absorb already digested food;
they lack the sort of skin we assume is on the outside of all creatures, since
they must soak up food through their surface. They don’t have a serious
nervous system or muscles. Rather than being a single organism, a tapeworm is
more like a giant colony of parts, each capable of independent activity.”
Interestingly, the U.S. Patent Office Manual, in sections pertaining to the rules for deposits of “biological materials” for patent purposes [2543], distinguishes nonparasitic and parasitic replicators: “Biological material includes material that is capable of self-replication either directly or indirectly. Direct self-replication includes those situations where the biological material reproduces by itself. Representative examples include bacteria, fungi including yeast, algae, protozoa, eukaryotic cells, cell lines, hybridomas, plant tissue cells, lichens and seeds. Indirect self-replication is meant to include those situations where the biological material is only capable of replication when another self-replicating biological material is present. Self-replication after insertion in a host is one example of indirect self-replication. Examples of indirect replicating biological materials include viruses, phages, plasmids, symbionts, and replication defective cells.”
“Replication processes” may include assembly or fabrication processes, where fabrication is the construction of parts and assembly is the rearrangement of parts. All kinematic replicators must engage in either fabrication or assembly activities, or possibly both. But not all replicators must engage in fabrication (e.g., when parts are externally provided, thus requiring only assembly activities) and not all replicators must engage in assembly (e.g., when the replicator has a unitary design (see D10) with no individual parts, thus requiring only fabrication activities).
I4. Replication Process Centralization.
Are replication processes concentrated in a single entity or subsystem, or instead
distributed to multiple entities or subsystems? In highly decentralized replication,
swarm construction [2544] may be used to
build “complex, composable structures” as is demonstrated by termites
and other social insects. See also D13.
I5. Replication Process Specialization.
Mill-style assembly processes involve highly specialized and purely repetitive
operations which produce standard building blocks, whereas manipulator-style
assembly processes involve more generalized “programmable operations that
can stack these building blocks to make a wide variety of products.” [208]
I6. Replication Process Timing.
When does the replicator replicate, and when does it do nonreplicative activities,
during its life-cycle?
I7. Replication Goal Specialization.
Despite the well-known human preoccupation with sexual matters, human physiology
devotes surprisingly little mass directly to the organs of reproduction. Specifically,
the 70,000 gm reference male body has only 40 gm of testes and ~60 gm of flaccid
penis, totaling only ~0.14% of body mass. The nongravid 58,000 gm reference
female body has 35 gm of uterus, 8 gm of ovaries, and perhaps ~50 gm of vagina,
again only ~0.16% of total body mass (Nanomedicine
[228], Table
8.9). In other words, >99.8% of human body mass does not directly execute
organismic replicative processes. Human tissue cells are slightly more specialized
to the goal of cell reproduction, with ~3.4% of tissue cell mass devoted to
the nucleus (Nanomedicine
[228], Table
8.17). Functional proteomics [2545]
indicates that Caenorhabditis elegans uses 6% of its open reading frames
to encode proteins required for cell division.
Of course, Dawkins would argue that all of an organism’s mass executes replicative processes in one way or another, and that the above direct/indirect dichotomy is artificial. For example, during pregnancy a woman’s body may raid itself for necessary minerals [2546], hence the medically unsubstantiated [2547] old adage and myth that a woman loses “a tooth for every child” [2548]. (In reality, calcium to build fetal teeth, when in short supply from the maternal diet, is absorbed from the mother’s bones, not from her teeth [2549]. Although orthodontic tooth movements are slightly greater during pregnancy [2550], apparently there is no direct mechanism for the physiologic withdrawal of calcium from teeth as there is from bone, so a developing fetus cannot calcify at the expense of the mother’s teeth [2551].) But metabolic processes that withdraw nutrients are not unique to pregnancy and hence cannot logically be ascribed to replicative processes per se – the human body frequently raids its stores of specific nutrients (e.g., glucose, fat, iron, etc.) during times of physiological stress or unusually high demand.
I8. Replication Process Subunit Assistance.
At what point during its construction can a daughter replicator begin assisting
in the construction of other replicators? See also F10.
I9. Replicator Parentage.
How many replicators cooperate in the building of a daughter device? For example,
Parham [2552] suggests 6-8 parent devices
might be useful, though most proposals and most natural systems typically invoke
only one or two active parental replicators.
I10. Replication Process Intermediaries.
An example in biology is the life cycle of slime molds, in which many originally
independent cells must come together to form the slug and then the fruiting
body, which then make many spores which become independent cells [2412].
Using replicators to build a robot factory which then builds the original replicator
was an early strategy first quantitatively explored by Freitas [1014]
in 1980. Sayama [9] proposes enhancing the robustness
of self-replication processes “by introducing an additional subsystem
that constructs a workplace prior to automaton construction.” (See also
C8 above.)
I11. Replication Auxeticity.
During the nineteenth century, the discovery that cells reproduce themselves
by dividing into two illuminated the origin of cells and became a cornerstone
of the cell theory [2553-2555].
The fission replication model is the intermediate case between the extremes
of a purely auxetic or factory replicator and a purely non-auxetic or unit replicator,
a very important and fundamental dimension in the replicator design space. Besides
individual cells, some multicellular animals also reproduce by fission. For
example, upon reaching a certain size a growing planarian pulls itself in two,
after which the head grows a new tail and vice versa, resulting in two smaller
individual animals [679].
I12. Self-Formation Modality.
According to Janusonis [784-786],
in self-formation the interaction between the forming object and the chaotic
medium (the environment or substrate) is controlled by the forming object’s
structure, and the structure is changed by the interaction, with the primary
object increasing in complexity. Three kinds of self-formation can be used in
manufacturing: self-alignment, development, and replication.
I13. Replication Process Parallelicity.
I14. Replication Process Digitality.
Replication based on DNA complementarity is considered digital, whereas autocatalytic
cycles are frequently processive (e.g., Section 5.1.4(2)).
I15. Replicator Parts Synthesis.
Parts may be fabricated by additive processes, subtractive processes, or by
other processes that alter the part without changing the quantity of physical
material that is present.
I16. Number of Distinct Onboard
Transformative Processes for Environmentally Available Parts. Edmonds
[502] discusses the quantity and variety of
transformative operation types in different classes of replicating systems.
I17. Ingestor Modality.
Physical inputs might not need to cross the replicator boundary if all assembly
processes take place external to the replicator, or if the replicator begins
operations with all raw materials already contained within its physical boundaries.
I18. Assembly Process Style.
Drexler [208] notes that there are at least
two different assembly styles. The first style he calls “construction-style
assembly” in which many small devices are used to work on or in a large
structure, analogous to constructing a building. Many assembly devices can work
in parallel to build up a surface, and the use of larger building blocks can
dramatically speed the rate of assembly. Construction-style products can be
assembled in poorly controlled environments (i.e., solvent bath), “simplifying
problems of heat and mass transport, and of environmental control....Alternatively,
eutactic environments of almost any desired size can be constructed by expanding
a gas-tight barrier.” [208] The second
style Drexler calls “manufacturing-style assembly” in which parts
and workpieces are manipulated and transported within larger mechanisms –
for example, convergent assembly sequences (Section 5.9.4)
“in which each structure is built from components within an order of magnitude
or so of the structure’s own linear dimensions. The motion of components
traces a tree in space: the trunk corresponds to the path traced by the final
workpiece as the final components are assembled, the branches correspond to
the paths traced by those components as they are assembled, and so forth. Convergent
assembly can be distributed in space in a manner that (1) provides an assembly
volume proportional in size to the workpiece at each stage, and (2) requires
only short-range transportation of parts between stages.” [208]
I20. Audit Trails. Does
the replicator log important events (e.g., replication) and register any new
replicator it makes with some external agency? Crude analogs in biology are
the homeostatic copy number control systems found in plasmids (Section
4.3.5).
I21. Process Accounting.
Does the replicator require payment of appropriate fees (or analogous transactions)
when an item is manufactured (e.g., does it have Digital Rights Management (DRM)
[2556] capabilities)?
I22. Process Security.
Does the replicator have effective limits on what it will manufacture? Most
importantly, will the replicator only accept and manufacture blueprints that
have been digitally signed by some external agency? Or will it manufacture any
blueprint you care to download (including illegal drugs, weapons, etc.)?
I23. Tamper Resistance.
Will the replicator resist attempts to physically subvert its security? Does
it possess active means (e.g., thermite to insure self-destruction if attacked)
to prevent physical subversion of its capabilities? Valuable commercial programs
commonly employ either primarily hardware-based locks, such as dongles, or primarily
software-based locks, such as authorization codes, to prevent unauthorized usage.
(Most hardware-based tamper resistant systems include some software, and vice
versa; the issue here is the relative emphasis.) Consider a replicator design
which incorporates, say, three major independent architectural means of preventing
unwanted replication (e.g., encrypted instruction sets, rare nutrients and broadcast
power) along with a nonevolvable design. For such a system, notes Yudkowsky
[2452], “by far the most probable
source of serious problems is human abuse, not a point mutation that somehow
gets past all three barriers. If a design is resistant to corruption by the
space of attacks employed by a human cracker, it will probably be resistant
to the space of probable point mutations.”
I24. Reliability of Manufacturing
Operations. The normal functioning of the ribosome produces one manufacturing
error in 1,000 to 10,000 operations, which limits the size of modules that can
be made without some form of error correction mechanism. (Section
4.2; see also L4 “Replication Fidelity”).
Errors in fabricated parts are of reduced concern if the parts are to be used
in an error-tolerant application or in a noncritical location of a component.
For instance, a protein with 1-2 randomly misplaced residues will probably still
fold correctly, and some nanomachine parts may work correctly with 1-2 random
atoms out of place; but 1-2 misplaced residues in the active site of an enzyme
can seriously degrade activity, and 1-2 misplaced atoms at a sliding interface
or in a binding site could utterly ruin the performance of nanomachinery.
I25. Error Correction Mechanisms.
Does the replicator include onboard error correction mechanisms (regardless
of whether instructions are stored onboard)?
I26. Offspring Separability.
Do offspring physically separate from their siblings after they are produced?
Lohn and Reggia [371] cite the ability of
distinct copies eventually to separate, after being initially adjacent, as a
criterion for self-replication in the context of cellular-automata-specific
replication. This factor is related to the multicellularity dimension F1.
Last updated on 1 August 2005