Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
4.3.3 Viruses
Viruses are acellular bioactive parasites that attack virtually every form of cellular life [228] and which cannot replicate except in the presence of a biological host cell. Viruses have diameters ranging from 16-300 nm [1751] – for example, poliomyelitis ~18 nm, yellow fever ~25 nm, influenza ~100 nm, herpes simplex and rabies ~125 nm, and psittacosis ~275 nm [1752]. Their shape is either pseudospherical with icosahedral symmetry, as in the poliomyelitis virus, or rodlike, as in the tobacco mosaic virus (TMV). Viruses consist of a core of RNA (most plant viruses and animal viruses such as the rhinoviruses, polio and flu viruses, and all retroviruses) or DNA (most bacterial and some animal viruses), but never both. This nuclear material is surrounded by a protein coat called a capsid, a quasi-symmetrical structure assembled from one or only a few protein subunits called capsomeres. A virus surrounded only by capsid is a naked virus; some viruses acquire a lipid membrane envelope from their host cell upon release, and are then called enveloped viruses. Attached to the capsid are other protein structures necessary for host infection, especially attachment or docking proteins and most importantly the ATP-powered DNA packaging motor (Section 4.5) that stuffs viral genetic material into the viral capsid [1753-1755]. The interior capsid volume is usually only slightly larger, and never more than twice as large, as the volume of the enclosed nuclear material [1693]. For instance, the adenovirus (one family of viruses that causes the common cold in humans) is an icosahedral particle ~70 nm in diameter (~180,000 nm3) containing one double-stranded DNA molecule ~11 microns (~35,000 bases or ~107 daltons) in length (~15,000 nm3), giving a composition of approximately 92% protein and 8% DNA by volume. By comparison, the TMV has 5% nucleic acid, the bushy stunt virus 17%, and the tobacco ringspot virus has 40% [1752].
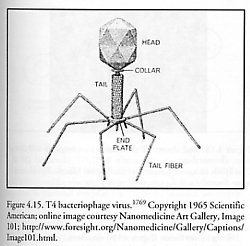
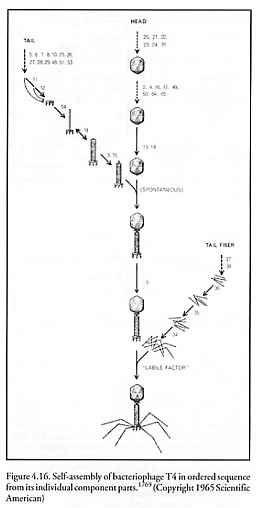
Bacteriophage viruses, first employed therapeutically against bacteria by d’Herelle in 1922 [1756], have recently been used as self-replicating pharmaceutical agents [1757]. One of the best-studied examples of the bacteriophages is the T4 virus (Figure 4.15). Like all viruses, T4 replicates primarily by self-assembly [1758-1762] in the final stage, albeit with the assistance of co-opted host-cell ribosomes (Section 4.2) – which are positional-assembly style devices – to make the “parts”. (Self-assembly and positional assembly are distinguished in the introduction to Chapter 4). The syringelike, 100-nm long, 20-nm wide cylindrical T4 tail assembly, made of 15 different proteins joined in 24 annular segments with an 8-nm inside bore, has a set of small fibers near the tip that attach to the plasma membrane of the host cell. After a lysozyme-like enzyme opens a breach in the host cell wall, the tail sheath thickens and contracts [1763], inserting a ~2.8 megadalton hollow core protein nanotube (80 nm long, 7 nm wide, 2.5 nm inside bore) through the host cell integument. The protein nanotube is then uncorked in response to chemical signals, and large-molecule (mostly putrescine and spermadine) pressurization of ~30 atm ejects a single 70-micron long, 2-nm wide DNA thread (~50% of T4 head volume) through the 2.5-nm nanotube aperture and out into the cell typically in ~3 seconds, a mean flow velocity of ~23 microns/sec [1764]. (Phage DNA is mechanically packed into capsids by molecular motors [1753].)* The double-stranded DNA thread rotates ~4000 times around its axis as it emerges [1765]. Under optimum conditions, initial injection velocity may start as high as 360 micron/sec with a minimum injection time of 0.23 sec [1764]. Once inside the host cell, the virus (specifically, its genetic material) uses the cell’s machinery, including some of the cell’s enzymes, to generate virus parts and the 168 kilobase T4 genome [1766], which self-assemble into new T4 virus particles (Figure 4.16). The phage head, tail, and tail fibers are assembled via independent pathways, and then are joined to form the complete virus [1767]. A single Escherichia coli bacterium injected with a single T4 phage virion at 37 oC in rich media lyses after 25-30 minutes (the first completed virus particle, est. mass ~0.3 fg, materializes ~13 minutes post-infection [1758]), releasing 100-200 phage particles that have replicated themselves inside the cell [1768].**
* Here we are describing only one well-known process of bacterial-assisted replication of an extensively studied bacterial virus. R. Bradbury notes that there are human viruses such as CMV and HSV that contain more than an order of magnitude more DNA and have a correspondingly more complex life cycle and assembly process.
** This describes only the lytic cycle of viral reproduction. In the alternative lysogenic cycle of some bacteriophage viruses, the phage’s DNA inserts itself into the bacterial chromosome, becoming a “prophage.” A host cell that carries a prophage has the potential to lyse, thus is called a lysogenic cell.
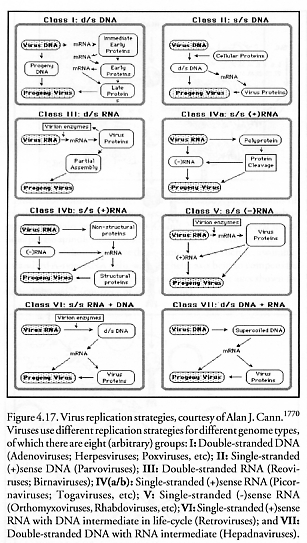
Thus the general logic of viral replication (Figure 4.17) appears to be to impose the exogenous viral “data tape” upon the cellular manufacturing machinery. This captive machinery fabricates all viral parts more or less simultaneously, whereupon these parts then self-organize by making use of prearranged conformational switching (Section 4.1.5) of the physical structure of parts and subassemblies as they self-assemble into new complete virion particles.
Last updated on 1 August 2005
{kind=link}
{kind=link}
{kind=link}