Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
4.2 Ribosomes: Molecular Positional Assembly for Self-Replication
In 1981, Drexler [197] pointed out that complex devices resembling biomolecular motors, actuators, bearings, and structural components could be combined to build versatile molecular machine systems analogous to machine systems in the macroscopic world, and then concluded: “Development of the ability to design protein molecules will open a path to the fabrication of devices to complex atomic specifications, thus sidestepping obstacles facing conventional microtechnology. This path will involve construction of molecular machinery able to position reactive groups to atomic precision.”
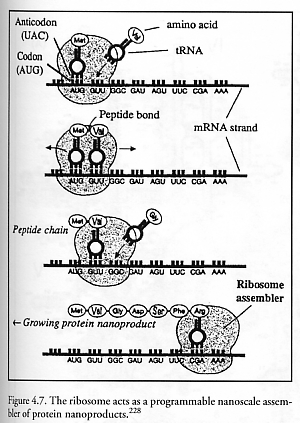
The best-known biological example of such molecular machinery is the ribosome, the only programmable nanoscale positional assembler currently in existence.* Numbering ~106-107 units per living eukaryotic cell and ~20,000 units per prokaryotic (bacterial) cell [1693], ribosomes act as general-purpose factories building diverse varieties of proteins by bonding amino acids together in precise sequences under instructions provided by a strand of messenger RNA (mRNA) copied from the host DNA (Figure 4.7), with energy for ribosome movement along the mRNA tape provided by the hydrolysis of guanosine triphosphate (GTP) [1693]. (The loading of transfer RNAs (tRNAs) with new amino acids by tRNA synthetase is powered by the conversion of ATP to AMP [1680].) Each ribosome is a compact ribonucleoprotein machine consisting of two subunits that fit snugly together, with each subunit consisting of numerous proteins associated with a long RNA molecule (rRNA). In mammalian cells, ribosomes are ~25 nm in diameter. The complete mammalian 80S ribosome unit has a total mass of 4.2 million daltons, including 2.8 million daltons for the large (~23 nm diameter) ribonucleoprotein 60S subunit containing 4998 bases and 49 proteins, and 1.4 million daltons for the small (~9 nm diameter) ribonucleoprotein 40S subunit containing 1874 bases and 33 proteins [1693]. The 80S unit is 60% rRNA by mass [1693]. Unlike other cellular polymerases, the ribosomal mechanism of action appears to depend almost entirely on the rRNA component; that is, ribosomes are giant ribozymes [1681-1684].
* There are three distinct types [1687] of ribosome – eubacterial, archaebacterial and eukaryotic – and multiple subvarieties among the ~108 different species on Earth.
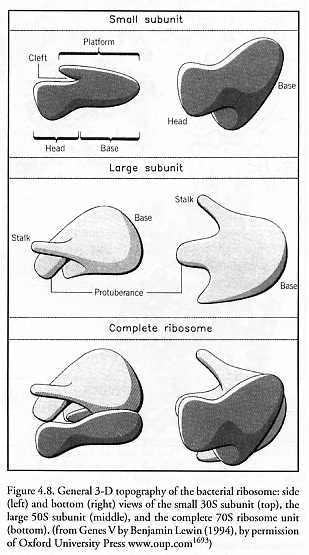
Bacterial ribosomes (Figure 4.8) measuring ~20 nm in diameter have been the most extensively investigated [1688]. The complete bacterial 70S ribosome unit is 66% rRNA by mass and is formed by the association of the large 50S subunit and the small 30S subunit through a network of intermolecular bridges [1689]. The large 50S subunit contains one 23S rRNA molecule composed of 2904 nucleotide bases and one 5S rRNA molecule composed of 120 bases, both embedded in a framework of 31 structural proteins which are located mainly at the periphery. The small 30S subunit contains one 16S rRNA molecule composed of 1541 bases embedded in a framework of 21 similarly mostly peripheral structural proteins [1690-1693].
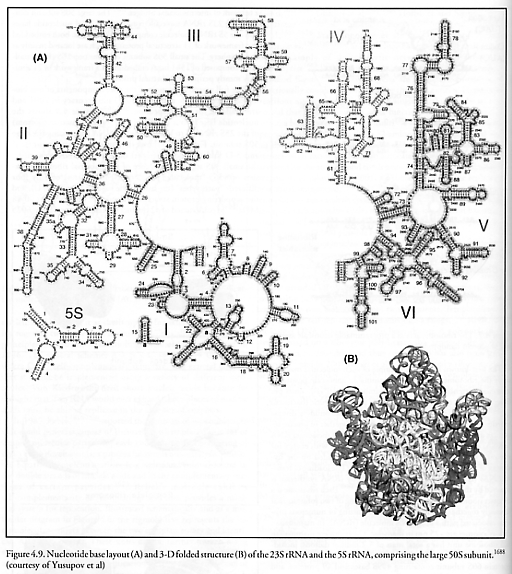
The full crystal structures of the core components of this biological machine were mapped to 0.55 nm accuracy in 2001 and are available for our inspection [1688, 1696]. Figure 4.9 shows the nucleotide base layout (top) and 3-D folded structure (bottom) of the 23S rRNA (I to VI) and the 5S rRNA comprising the large 50S subunit. Figure 4.10 shows the nucleotide base layout (left) and 3-D folded structure (right) of the 16S rRNA comprising the small 30S subunit. The core of the bacterial ribosome consists of 4565 “molecular parts” (nucleotide bases) of only four different “parts types” (the nucleotide bases A, C, G, and U), and the supporting protein structural framework of total mass ~0.85 MDa [1693] is constructed of 52 distinct proteins composed of ~6025 amino acid residue “parts” (assuming ~0.1411 kD/residue [1680]) of 20 different “parts types.” Thus we may say that the bacterial ribosome assembler is a partially self-replicating machine that is built of ~10,590 “molecular parts” and 24 different “parts types.” (Using the same counting method, the mammalian ribosome has ~18,780 parts.) These totals do not count the external supporting environment which includes tRNA “parts holders” and aminoacyl-tRNA synthetase “parts holder reloaders” of 20 different types each, various elongation factors that cyclically associate and dissociate with the ribosome, rRNA polymerase enzymes that construct both mRNA “instruction tapes” and rRNA feedstock molecules via transcription, various chaperone molecules to assist in proper protein folding, and the mRNA instruction tape itself. Note that because the ribosome cannot produce its own most important component, rRNA, the ribosome is not a fully self-contained self-replicating machine, but may most precisely be regarded as the key component of a self-replicating system.
During assembly operations (aka. “translation”), the space between the subunits is largely occupied by tRNAs (measuring ~6 nm wide x ~6 nm tall), whose anticodons base pair with mRNA codons in the small 30S subunit. The 3’-CCA end of each tRNA “parts holder” molecule, which carries the growing polypeptide chain and grasps the incoming amino acid “molecular part,” reaches into the large 50S subunit – the location of the peptidyl transferase center where peptide bond formation is catalyzed. In bacteria at 37 oC, 15-40 amino acids per second are added to the growing polypeptide chain [1693-1695] with an overall error rate of ~0.05% per codon that is largely due to mistaken tRNA recognition [1693]. If the bacterial ribosome’s ~4.2 x 10-21 kg mass consisted entirely of proteins, then this simple biological assembler could (very loosely) be said to have a self-replication time of order 270-710 sec – the time required to synthesize its own mass, in proteins. In eukaryotes the rate of protein synthesis is often lower, sometimes as slow as ~2 residues/sec [1693].
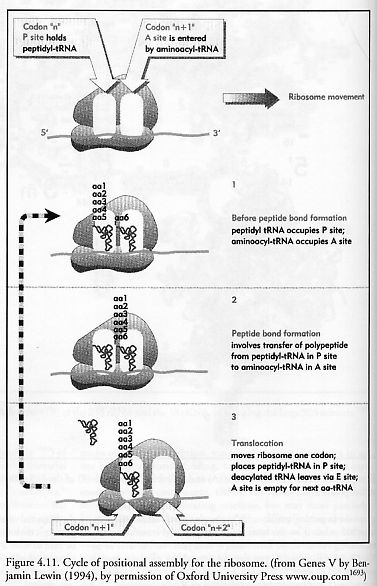
How does the ribosome molecular assembler work? Many details are now known [1683-1686], but we have space here to offer only the most cursory summary. The ribosome has within its physical structure three active stations – the A site, the P site, and the E site – through which all tRNAs must pass, in sequence (A → P → E), during translation [1693]:
The A site (or entry site), is the only site that can be entered by an incoming tRNA with an amino acid attached to it, called an aminoacyl-tRNA. The particular desired tRNA (of 20 possible types, one for each amino acid type) must be discriminated from all its tRNA brethren along with whatever other solutes can fit into the A-site pocket using codons with only three base pairs for discrimination. (In fact, most codons use only two of the three base pairs for recognition, so some tRNAs are recognized by as few as 4 hydrogen bonds [1697].) At the start of translation, the ribosome positions itself over the mRNA tape so as to expose to the A site the codon on the tape that represents the next amino acid due to be added to the chain.
The P site (or donor site) lies over the mRNA codon representing the most recent amino acid to have been added to the growing polypeptide chain. This site is occupied by a tRNA to which the growing polypeptide chain is attached, called a peptidyl-tRNA.
The E site (or exit site) holds a tRNA from which all peptide and amino acid has been removed, called a deacylated tRNA. While the E site is occupied, the affinity of the A site for aminoacyl-tRNA is greatly reduced, preventing the next aminoacyl-tRNA from entering before the ribosome is ready. This site is transiently occupied by the tRNA en route between leaving the P site and being released from the ribosome back into the cytosol. Once released, an appropriate aminoacyl-tRNA synthetase enzyme attaches to the deacylated tRNA [1698], adds the appropriate amino acid, then detaches, leaving behind a refreshed aminoacyl-tRNA that is ready to cycle again.
Figure 4.11 shows schematically the addition of a sixth amino acid (aa6) to a growing peptide chain that is already 5 residues long. During the next translation cycle the new aminoacyl-tRNA bearing the amino acid “aa6” enters the A site and is secured. Next, the existing polypeptide is transferred from the peptidyl-tRNA in the P site to the aminoacyl-tRNA in the A site, forging a new peptide bond (via peptidyl transferase activity on 23S [1681]) between aa5 and aa6. The deacylated tRNA moves from the P site to the E site; the ribosome translocates one codon to the right (~1.3 nm [1699]) along the mRNA tape, moving the new peptidyl-tRNA from the A site to the P site; the deacylated tRNA is ejected from the E site; and the A site is now empty and ready to accept the next aminoacyl-tRNA. Observe that all peptide “parts” and their tRNA “parts holders” (analogous to fixture-based replication; Section 5.1.8) are geometrically confined to specific locations during all key steps in translation, and chemical reactions that form covalent bonds are restricted to a specific site and are digitally sequenced. The ribosome – in effect a digitally-controlled mechanical ratchet [1686] in which peptide bond formation is driven by the codon-anticodon interaction – exhibits true positional assembly of molecular parts. There is even a special mechanism (tmRNA [1700]) to unjam and restart ribosomes that have become stalled on broken mRNA tapes.
Interestingly, the ribosome itself is assembled via self-assembly in a series of sequential reactions in which groups of proteins (built by ribosomes) associate with rRNA (made by RNA polymerases I and III [1701-1703], themselves 500-600 kD protein-based enzymes [1680] built by ribosomes), and the structure then folds so that the next group of proteins can join [1704-1706]. Hence the ribosome requires both self-assembly and positional assembly in order to achieve its own self-replication. (In a different context, “mechanically assembled monolayers” or MAMs have been fabricated by combining the self-assembly of surface grafting molecules with mechanical manipulation of the grafting points in the underlying elastic surface [1707].)
Other bioassembler units such as polyketide synthases and nonribosomal peptide synthetases [1708-1710] also exist and are being engineered [1711-1715], but these enzymes generally only make one type of product per assembly. Loosely speaking, some bacteria produce polyketides in a Detroit-like assembly-line fashion [1718-1722]. These microbes contain dozens of modular enzymes, each of which assembles one component of the polyketide and then passes it along to the next enzyme in the sequence, eventually resulting in the complete synthesis. These modular synthase chains can be engineered [1715-1717].
Last updated on 1 August 2005
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}