Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
4.5.1 Positional Assembly Using DNA
Starting in 1994, Niemeyer [1451] and Smith [1417] exploited DNA specificity to generate regular protein arrays, using “oligonucleotide-directed self-assembly of proteins” [1451] and “nucleoprotein-based nanoscale assembly” [1417]. Shi and Bergstrom [1960] attached DNA single strands to rigid organic linkers, showing that cyclical forms of various sizes could be formed with these molecules. Working from the insight that DNA could serve as an assembly jig in solution phase, Smith and Krummenacker [1961] devised a possible method for the nanoassembly and covalent linkage of protein “nanoparts” into specific orientations and arrangements as determined by the hybridization of DNA attached to the proteins, called DNA-Guided Assembly of Proteins (DGAP) [1962]. In this method, multiple DNA sequences would be attached to specific positions on the surface of each protein, and complementary sequences would bind, forcing protein building blocks (possibly including biomolecular motors, structural protein fibers, antibodies, enzymes, or other existing functional proteins) together in specific desired combinations and configurations, which would then be stabilized by covalent interprotein linkages. This technique could also be applied to non-protein components that can be functionalized at multiple sites with site-specific DNA sequences, although proteins, at least initially, may be more convenient building blocks due to their size, their surface chemistry, the wide variety of functions and mechanical properties they can confer on the resulting assemblies, and the many existing techniques for introducing designed or artificially evolved modifications into natural proteins of known structure. In 2001 a nascent company, Molecubotics, Inc. [1963], put forward a business plan to pursue the DGAP method of molecular assembly [1964] and to begin seeking funding.
Others are exploring methods for: covalently attaching functional (fusion) proteins to a DNA backbone in a specified manner at ~8.5 nm (25 base-pair) intervals [1417]; addressable protein targeting in macromolecular (e.g., nucleoprotein) assemblies [1450-1457]; attaching specific DNA tag sequences to protein molecules such as enzymes to a specific region on the surface of the target protein well away from the active site [1965]; laying of silver wire [1966, 2019], gold wire [1967-1969], and other nanowires [1970-1974] using DNA as a molecular scaffolding or possible template for circuit construction [1975], including metal-centered DNA lattices [1976], metal nanoparticle assemblies [1977, 1978], and nanotube localization [1979]; decorating DNA with fullerenes [1980] and dendrimers [1414]; assembling DNA-tagged nanometer-sized gold [1981, 1982], magnetic [1983], or other particles into larger clusters, or into spontaneous lattices [788] or magnetic chains [1984]; silicon component aggregation on DNA-directed 2-D array [1985] and “protein stitchery” [1986]. Braun et al [1987] have used DNA scaffolding with several specific proteins as a template for the self-assembly of carbon nanotube based transistors, with metallic nanowires at either end. DNA self-assembly can sort mixed nanotubes into the metallic and semiconducting varieties [1988]. Cook et al [1989] show how DNA tiles can be used for self-assembled electronic circuit patterns and as a scaffold for functional devices such as molecular electronic circuits, and that several circuit patterns – such as demultiplexors, random-access memory, and Hadamard matrix transforms – can, in principle, be self-assembled from a small number of DNA tile types.
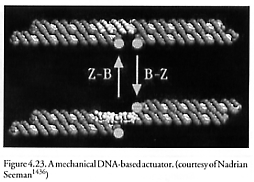
Seeman’s group [1434-1450] at New York University has reported the design and construction of 2-dimensional DNA arrays (which might serve as templates for nanomechanical assembly) [1437, 1441, 1442, 1990] and a mechanical DNA-based actuator [1436] (based on the solution-ion-based B-Z structural transition in DNA; Figure 4.23) and similarly actuated rotary motors [1434] and a bipedal DNA nanowalker with 36 base-pair-long legs [2342]. Seeman is now collaborating [1991] with computational chemists to achieve “the design and fabrication of practical nanoscale devices” and “to make rapid progress in demonstrating DNA based nanoscale devices.”* Erik Winfree at CalTech is investigating using DNA tiles to perform computing tasks [1989, 1992-1997].
* In a lecture at the 10th Foresight Conference in Washington DC in October 2002, Seeman noted that “self-replicating systems” are explicitly being studied by his research group, but that he “didn’t have time for this today, the work is on hold for now.” A lecture slide entitled “Challenges for structural DNA nanotechnology” included the item: “To produce systems capable of self-replication – economy; evolvability.” Seeman has already built DNA-based nanomechanical devices [1434, 1436] including “sequence-dependent devices [that] can provide the diversity of structures necessary for nanorobotics”[1998], and in 2002 Seeman wrote [1999]: “The development of self-replicating systems using branched DNA appears today to be somewhat oblique [1450], but it nevertheless represents an exciting challenge that will significantly economize on the preparation of these systems and enhance their evolvability. Ultimately, structural DNA nanotechnology must advance from the biokleptic to the biomimetic, not just using the central molecules of life, but improving on them, without losing the inherent power they possess.” According to Seeman in late 2003 [2000], the replication scheme, first presented in 1991 [1450], involves DNA knot topologies [2001] and requires the ability to characterize what is synthesized via direct structural observation, rather than by gel or other biochemical techniques; forthcoming 3-D arrays “make these systems amenable to crystallographic characterization before picking up the knot work again and then applying it to replication.”
Other DNA-based actuation is known. For example, sequence-specific DNA hybridization is commonly used to bend silicon microcantilevers [2002-2006]. Alberti and Mergny [2010] synthesized a sequence-dependent DNA “piston” composed of a 21-base oligonucleotide that displays a 5-nm, two-stroke, linear motor type movement. Li and Tan [2011] have made a single-DNA-molecule inchworm motor. Shu and Guo [1754] synthesized a 30-nm long chimeric pRNA (DNA-packaging) motor made from six strands of RNA surrounding a center strand of DNA: in the presence of ATP, the RNA strands push the DNA axle in succession, spinning it around and producing 50-60 pN of force.
Researchers from Ludwig Maximilians University in Germany have built a simple molecular machine from DNA that can bind to and release single molecules of a specific type of protein [3110]. This “DNA hand” can be made to select any of many types of proteins, and could eventually be used to construct materials or machines molecule-by-molecule. “The researchers used DNA branch migration, a method that allows a DNA nanostructure to switch between several arrangements of its parts, to construct the DNA hand. In one configuration, the structure contains an open sequence of bases that binds to a specific protein, and so can grab that type of protein. A second configuration does not contain the open sequence, and so drops the protein. The rearrangements are reversible, allowing the tiny machine to repeatedly grab and drop a molecule of a specific type of protein. DNA aptamers, or strands that bind to specific molecules, can be selected from a pool of DNA sequences, making it possible to construct a DNA hand that binds to any type of protein.” [3110] The DNA hand was demonstrated by having it repeatedly grab and drop molecules of the protein thrombin.
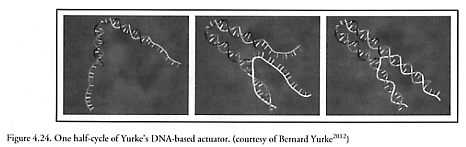
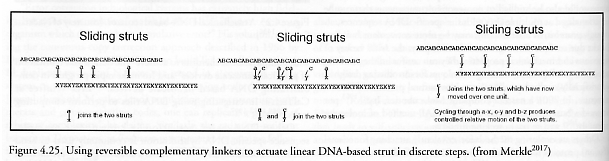
Bernard Yurke at Bell Laboratories and Andrew Turberfield at the University of Oxford have synthesized a sequence-dependent DNA-based actuator which has been cycled 7 times consecutively by inserting and removing a special “fuel” strand, causing the “nanotweezer” to close from a ~7 nm separation to a ~1 nm separation (Figure 4.24), taking ~13 sec per cycle [2012-2015]. (More recent work [2016] has focused on a continuously running DNA nanomotor.) Merkle [2017] has proposed a similar chemically-driven DNA-based “sliding strut” actuator (Figure 4.25) which demonstrates that biotechnology and self-assembly can be used to make positional devices. Hao Yan and colleagues in Reif’s lab at Duke University in Durham, North Carolina have devised X-shaped DNA tiles that link up in a square grid with some of the strands consisting of sections of DNA that can lengthen and shorten by 6.8 nm like tiny pistons, making a net whose mesh size can expand or contract under chemical control [2018-2020]. RNA polymerase has been utilized as a 15-20 pN positioning motor that “can be incrementally ‘walked’ or positioned along DNA with nanometer scale precision.” [2021]
Reif [2022, 2023] has designed (but not yet built) “the first autonomous DNA nanomechanical devices that execute cycles of motion without external environmental changes. These DNA devices translate along a circular strand of ssDNA [single-stranded DNA] and rotate simultaneously.” Reif’s first DNA device, called “walking DNA,” would achieve random bidirectional motion around a circular ssDNA strand by using DNA ligase and two restriction enzymes that consume ATP as their energy source. Reif’s second DNA device, called “rolling DNA,” would achieve random bidirectional motion by using DNA hybridization energy in trapped states with no DNA ligase or restriction enzymes. The emerging field of DNA-based molecular construction appears quite active [2024].
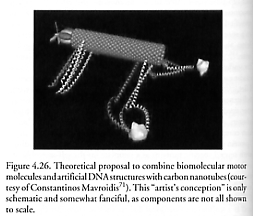
Mirkin’s group [2025] has created 2-D and 3-D architectures with DNA and inorganic nanoscale building blocks, including using massively parallel tip arrays to write patterns using a DNA-based “ink” [2026]. Mavroidis’ group [71] is studying combining artificial DNA structures with carbon nanotubes (Figure 4.26). Jaeger and colleagues are investigating 9-13 nm self-assembling tectoRNA geometrical figures [1675-1678], and von Kiedrowski’s group [1429] has used synthetic three-armed tris-oligonucleotidyl building blocks to assemble 3-D DNA nanoscaffolds such as supramolecular tetrahedra. Others [2052] have tailored allosteric responses in ribozymes – capabilities which might also be useful in the context of positional nanoparts assembly. There have also been numerous experiments with artificial nucleic acids [2027] including unnatural nucleobases [2028-2030] and nucleosides [2031-2033] (e.g., thiophene and furan heterocycles), “universal” bases [2034, 2035], peptide nucleic acids (PNA) [2036-2038], threose nucleic acid (TNA) [2039], and extended (unnatural) genetic alphabets [2040-2044].
Last updated on 1 August 2005
{kind=link}
{kind=link}
{kind=link}
{kind=link}