Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
4.3.7 Mitochondria
Mitochondria are energy-producing organelles present in virtually all eukaryotic cells. After the nucleus and the endoplasmic reticulum, the mitochondrion is often the largest organelle in most animal cells and is most likely an ancient endosymbiont [1818] of the eukaryotes. (Even secondary endosymbiosis, eukaryotes engulfing eukaryotes, has been found in cryptomonads [1819-1821] and chlorarachniophytes [1820-1822].) The typical time-averaged dimensions of the mitochondrion are roughly cylindrical, with a 0.5-1.0 micron diameter and a ~3 micron length (and rarely, up to 10 microns [1823]) – about the size of a bacterium (~1 µm3 or ~1.2 x 10-15 kg each). Except for plant chloroplasts, mitochondria are unique among organelles in having their own DNA genomes, ribosomes and tRNAs that are quite different from those found in the cytoplasm. The interior of the mitochondrion is filled with a semifluid gel-like matrix that contains a concentrated mixture of hundreds of different enzymes related to aerobic cellular respiration and oxidation, plus several identical copies of the mitochondrial mtDNA genome which encodes the two RNA species present in mitochondrial ribosomes (12S and 16S rRNAs), a full set of transfer RNAs (22 genes) required for protein synthesis [1824], and 13 polypeptide constituents of respiratory chain complexes [1825]. The mitochondrial genome [1825, 1826] consists of a circular DNA molecule of ~11 million daltons, coding for about a dozen polypeptides. The human mtDNA molecule has 16,569 base pairs and a contour length of ~5 microns (cf. 16,831 base pairs for elephant mitochondria [1827], etc.).
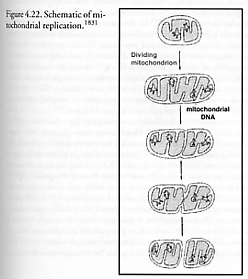
When cellular requirements for ATP increase, mitochondria pinch in half (i.e., binary fission; Figure 4.22) to increase their numbers, then both halves regrow to the former full size [1828]. (Doubling time for mitochondria in chloramphenicol-treated Tetrahymena pyriformis cells is ~2 hours [1829], and 2-5 hours in Physarum polycephalum cells under starvation conditions [1830].) It is a process very similar to the replication of prokaryotes, with DNA polymerase gamma processing the mtDNA, except that over evolutionary timescales many of the genes needed for independent replication have been offloaded to the much larger genome residing in the eukaryotic cell nucleus. Thus mitochondria are self-replicating organelles, but they require the assistance of cell-supplied proteins and lipids.
This illustrates yet another interesting variant in the logic of kinematic self-replication. Just as “inherently safe” (Section 5.11) replicators may be created by employing the “broadcast architecture” [208-210] wherein key information (instructions) essential for replication is exclusively provided from without, inherently safe replicators may also be created by employing a “vitamin architecture” in which key materials (physical components or parts) essential for replication are exclusively provided from without. In kinematic replicator theory, such key materials or components have often been called “vitamin parts” [2, 216, 271].
So, for example, if any of three specific outer membrane proteins – Mmm1p [1832], Mdm10p [1833], and Mdm12p [1834] – are removed from a mitochondrion, then fission is not possible and the result is one giant parent mitochondrion instead of two smaller daughter mitochondria. These three proteins may be considered as “vitamin parts” relative to mitochondrial replication. Similarly, most mitochondrial lipids, the vast majority of mitochondrial proteins (including DNA and RNA polymerases, ribosomal proteins, aminoacyl tRNA synthetases, etc. [1835, 1836]), and even the mitochondrial 5S rRNA [1837] are encoded by nuclear DNA and synthesized in the extramitochondrial cytosol, and thus must be imported into the mitochondrion as additional “vitamin parts” for replication to occur. (See also Sections 5.6 and 5.1.9 (A1, E4, I3).)
Last updated on 1 August 2005
{kind=link}