Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
5.1.6 Taylor Categorization of Reproducers (1999)
In his 1999 Ph.D. thesis on artificial life, Tim Taylor [444] focuses on three important dimensions defining the replication design space:
(1) Explicit-Implicit Coding scale – the degree to which the algorithm for reproduction (the way in which the process is specified and controlled) is explicitly encoded on the configuration being reproduced, rather than being implicit in the physical laws of the world. This distinction defines a continuum of possibilities.
(2) Auto-Assisted Reproduction scale – whether reproduction happens purely by the action of the physical laws of the world on the configuration to be reproduced (auto-reproduction), or whether it also requires auxiliary physical (or logical) machinery (assisted-reproduction). Taylor suggests that this distinction defines only a dichotomy, but allowing varying degrees of “assistance” would produce a continuum along this design dimension as well.
(3) Limited-Indefinite Hereditary scale – the number of different configurations that exist, connected by mutational pathways, that are capable of reproducing their specific form – that is, the distinction between limited hereditary reproducers and indefinite hereditary reproducers, a distinction also cited by Dawkins (Section 5.1.1) and Szathmary (Section 5.1.3). This factor could be expressed as the proportion of all possible mutations the reproducer may experience that will result in the production of distinct, yet viable, reproducers. This distinction also defines a continuum of possibilities, and indeed this axis may be infinitely long.
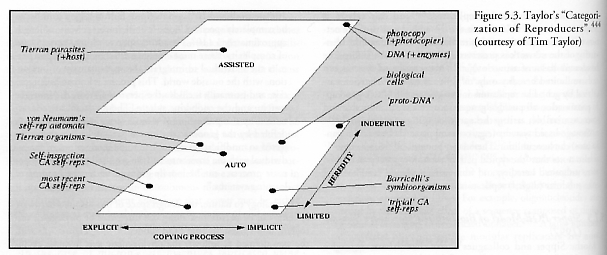
Taylor notes that these three scales seem “generally independent of each other” – i.e., they are mutually orthogonal dimensions in the design space – although he admits “the more explicitly encoded the reproduction algorithm is, the less likely, in general, it is to be an indefinite hereditary reproducer because of the increased chance of mutations disrupting the copying process.” Figure 5.3 shows Taylor’s qualitative categorization of some well-known replicating systems.
Taylor [444] notes that in his schema, Tierran organisms (Section 2.2.1) and von Neumann’s self-reproducing cellular automata (Section 2.1.3) “are placed midway along the limited-indefinite hereditary scale because, although both representations are capable of supporting universal computation in principle, only mutations which retain the ability to reproduce will be viable. In particular, in von Neumann’s architecture a mutation which affects a section of the tape which encodes the constructing automaton, the copying automaton or the control automaton will generally disrupt the ability of the combined automaton to produce viable offspring. Likewise, in Tierra a mutation which affects a section of the program which encodes the self-reproduction algorithm will generally disrupt the ability of the program to reproduce.”
Taylor [444] classifies DNA reproduction as assisted, because DNA can replicate “only with the aid of a host of enzymes to control the unwinding of the double helical structure, the polymerization of the individual bases of the new molecule, etc. At the same time, DNA is, in the presence of suitable enzymes, capable of indefinite heredity assisted-reproduction; the enzymes are able to copy any DNA double helix, no matter what sequence of bases it comprises. Within this context, the copying process is implicitly encoded in the DNA’s environment (in the enzymes which support the reproduction process, and in the physical laws governing the inherent bonding affinities of the bases) rather than being explicitly encoded upon individual strands on DNA. In contrast, a cell as a whole can be considered an auto-reproducer, as it can completely direct its own reproduction (in the presence of sufficient energy and raw materials from the environment), but its hereditary potential is slightly more restricted than DNA assisted-reproduction, because some mutations may disrupt the ability of the cell to reproduce. Similarly, each of Barricelli’s symbioorganisms (Section 2.2.1) can be considered, collectively, as an auto-reproducer, although individual digits within the symbioorganism are assisted-reproducers. The same analysis can be applied to collectively-autocatalytic reaction networks (e.g., Kauffman [2396]).”
The hypothetical ‘proto-DNA’ (i.e., a desirable seed for open-ended evolution) is placed in the auto-implicit-indefinite hereditary corner of the space because “the seed should be auto-reproducing (i.e., not rely upon auxiliary machinery) if it is to have a reasonable chance of spontaneously emerging, and it should be an indefinite hereditary reproducer to support an ongoing, open-ended evolutionary process. The requirement that it be an indefinite hereditary reproducer is most easily fulfilled if it reproduces implicitly.”
Last updated on 1 August 2005
{kind=link}