Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
4.3.6 Eukaryotic Cells
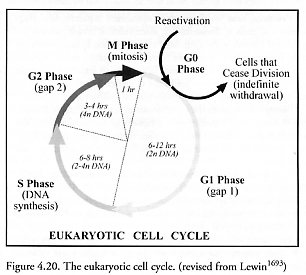
The replication of eukaryotic plant and animal cells (cells having a distinct nucleus and organelles) is a somewhat more complex process than for prokaryotes (cells having no distinct nucleus or membrane-enclosed organelles), following a tightly controlled cell cycle program [1797-1802] with well-defined phases (Figure 4.20). There are even checkpoints [1803] to verify that each phase has been successfully completed, and if not, the cell may be compelled to self-destruct (e.g., undergo apoptosis [1804]). Eukaryotes span a considerable size range. For example, most human cells measure between 2-120 microns in diameter [228].
Cells not currently replicating are said to be in the G0 phase, and begin the replicative process by entering the cell cycle at the first “gap” or G1 phase. G1 phase typically lasts 6-12 hours and is the first growth phase, during which time the cell increases its mass and both RNAs and proteins are synthesized, but there is no DNA replication. Following G1 (and after passing a commitment checkpoint) is the “synthetic” S phase, lasting 6-8 hours, wherein new DNA is synthesized and the existing chromosome set is duplicated [1805]. (Note that a few plant eukaryote reproductive cycles include complete organisms that are haploid, not diploid [1806].) The cell also continues to grow during S phase. All this activity depletes the cell’s energy resources, so a second period of growth (without DNA synthesis) follows, called the second “gap” or G2 phase, lasting 3-4 hours. Finally having rebuilt its physical resources (and after passing another commitment checkpoint), the cell now enters M phase or mitosis, lasting <1 hour, whereupon the nuclear material is separated and the parent cell divides into two daughter cells. Virtually all biochemical synthesis activities come to a halt during M phase [1807]. The daughter cells are born into G1 phase, and may either proceed through another replicative cell cycle or may exit the cycle and enter the noncycling G0 phase.
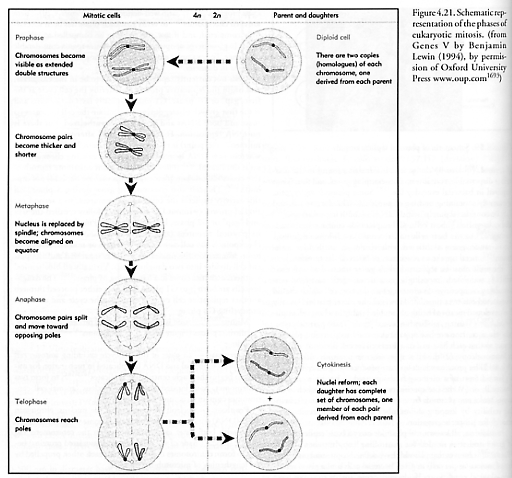
Mitosis [1808-1810] itself is a highly structured process involving several distinct phases (Figure 4.21) and checkpoints [1811, 1812] of its own, including:
Interphase – all parts of the cell cycle excluding mitosis; cell metabolism is normal and DNA is replicated in preparation for mitosis to begin; the single centrosome replicates [1813-1817] to form two centrosomes, both located outside the nucleus (animal cell centrosomes contain structures called centrioles).
Prophase – nucleoli disappear from the nucleus; chromatin fibers (DNA) condense, coil tightly, and become visible as chromosomes with both sister chromatids joined at the centromere; the mitotic spindle (made of microtubules and associated proteins) begins to form; centrosomes move away from each other, propelled by the lengthening of microtubular bundles.
Prometaphase – the nuclear membrane separates into fragments and then is disassembled; microtubules invade nuclear region and interact with condensed chromosomes; spindle fibers extend from each pole and move toward the equator; some microtubules connect to the kinetochore (a specialized attachment region on each chromatid which lies in the constricted region defined by the centromere) while other microtubules radiate to the equator.
Metaphase – with the centrosomes at
opposite ends of the cell, the chromosomes convene on the equatorial metaphase
plate (the middle region); chromosome centromeres are aligned and the identical
chromatids are attached to kinetochore tubules radiating from each pole.
Anaphase – paired centromeres of each
chromosome divide, liberating sister chromatids from each other; spindle fibers
shorten, pulling the newly synthesized chromosomes (DNA) to one end of the cell
and the original chromosomes to the other end of the cell; the cell begins to
pull apart.
Telophase – non-kinetochore fibers continue to elongate the cell; chromosomes begin to uncoil; daughter nuclei begin to form (opposite of prometaphase). A cleavage furrow forms and the two cells split apart, a process called cytokinesis. In animal-cell cytokinesis, a cleavage furrow appears, beginning as shallow groove in cell surface. On the cytoplasmic side of the furrow, a contractile ring of microfilaments is made from the protein actin; as the ring of microfilaments contracts, the cleavage furrow deepens until the parent cell is pinched apart into two daughter cells. In plant cells, a cell plate forms across the equator during telophase, whereupon vesicles from the Golgi body are driven along microtubules to the middle of the cell and the subsequent fusion of vesicles forms the membranes.
As with bacteria, the replicative logic for eukaryotes seems to be: Starting with one parent replicator, build enough of the constructor to enable tape copying, then replicate the instruction tape, then finishing building the rest of the constructor, which is composed of many specialized devices like a factory (e.g., ribosomes, mitochondria, Golgi membranes, etc.). Once there are two instruction tapes and enough factory devices to comprise two complete constructors, and a sufficient reserve of stored energy, the tapes and factory devices are distributed into two systems of roughly equal size having nearly identical sets of components. The two identical systems are then physically separated, making two independent daughter replicators.
Last updated on 1 August 2005
{kind=link}
{kind=link}