Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
4.3.5 Plasmids
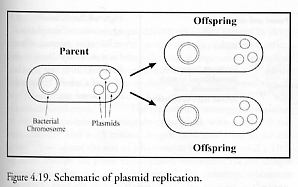
Plasmids [1782, 1783, 1793] are extrachromosomal elements found in most bacteria (and also in some eukaryotic cells). A plasmid is an autonomous circular DNA genome that may replicate along with the bacterial genome (Figure 4.19) or may be under a different type of control [1693]. Usually the genetic information found in the plasmid is dispensable, contains no housekeeping genes, and thus is not required in bacterial everyday life [1782]. Some plasmids are cryptic, apparently containing nothing of particular value for the cell, and may be considered purely “selfish DNA” – both in prokaryotic [1784] and eukaryotic [1785] host cells. Other plasmids confer selective advantage [1786] to the host cells under certain conditions, including, for instance, genes associated with virulence, antibiotics resistance [1787], metal uptake or resistance, or intercellular transfer.
Plasmids must be replicated, their genes transcribed and their mRNAs translated, imposing a parasitic energetic cost (and presumably a competitive disadvantage) on the host cell which should be selected out over time [1782]. However, the clever plasmid usually has developed means of being maintained stably in a host cell population [1788, 1795], using stability functions [1782] of: (1) the partition type, wherein a centromere-like apparatus is used to partition plasmid copies so that each bacterial daughter cell (at cell division) acquires at least one copy; (2) the killer type, wherein plasmids encode conditional killer proteins which only are produced when a cell has lost a plasmid (sort of a memory function used to punish a naughty host cell); or (3) the site-specific recombination type, wherein enzymes make two plasmids from one copy (which is a dimer), helping stability by keeping the number of plasmid molecules high enough for proper segregation.
In addition, all known real plasmids carry a basic replicon, a set of genes and sites needed for controlling copy number [1789-1793]. Wagner [1782] observes that plasmids have defined copy numbers (number of molecules per cell) in given bacterial cells under given sets of physiological conditions. If plasmids were unable to make head counts and correspondingly regulate their own replication (which is always carried out using host enzymes), their copy numbers would fluctuate because of natural physiological variations. After many generations of growth, this would result in bacterial populations in which some cells might contain as little as one plasmid at cell division while others could carry hundreds. Such a scenario is not beneficial for a plasmid, since in the first case, one of the daughter cells will have lost the plasmid, in the second case the plasmid load may kill the host cell. Hence, plasmids have evolved copy number control systems. Copy number control is homeostatic: a plasmid can “measure” its copy number in the cell, and use this information to affect the replication frequency at the plasmid origin of replication. Plasmids use different devices to accomplish this, including iterons (repeated DNA sequences) [1793, 1796] and antisense RNAs [1793-1795].
Last updated on 1 August 2005
{kind=link}